

分子植物科学卓越创新中心范敏锐研究组合作解析渗透压调控的胆碱转运蛋白BetT的结构与机制
文章来源:分子植物科学卓越创新中心 | 发布时间:2024-08-15 | 【打印】 【关闭】
2024年8月15日,中国科学院分子植物科学卓越创新中心范敏锐研究组与复旦大学张金儒团队、美国爱荷华州立大学Gwyn A. Beattie团队合作,于Science Advances杂志发表了题为“Structure and mechanism of the osmoregulated choline transporter BetT”的研究论文,解析了与细菌渗透压保护相关的胆碱转运蛋白BetT的高分辨率冷冻电镜结构,阐明了BetT识别胆碱的分子基础以及活性调控的自抑制机制,为改造植物提高叶绿体胆碱摄取能力以及抗逆能力提供了重要借鉴。
光合作用是植物生长发育的基础。在逆境条件下,植物通过一系列的策略保护自身免受光抑制损伤。大量研究表明甜菜碱对于保护植物在高盐、干旱、高温等环境压力下免受光抑制损伤具有重要作用,在多种植物和农作物中的研究表明引入甜菜碱合成相关基因可以有效地提高植物的耐逆能力和光合固碳效率。植物的甜菜碱在叶绿体中合成,需要首先将前体胆碱转运到叶绿体内,进一步生成甜菜碱。研究表明细菌中也存在类似的胆碱转运和甜菜碱合成途径,生成的甜菜碱可以帮助细菌抵御外界渗透压的变化,比如高盐环境。
本研究聚焦细菌中渗透压调控的胆碱转运蛋白BetT,利用冷冻电镜技术解析了来源于丁香假单胞菌的BetT(PsBetT)处于apo和胆碱结合状态的结构,分辨率分别为2.57埃和2.58埃。结构显示BetT蛋白采用经典的LeuT折叠模式,并形成domain-swapped的三聚体,其中每个单体的C端结构域与相邻单体的跨膜结构域相互作用。底物胆碱结合在由三个高度保守的色氨酸形成的特殊口袋中,主要涉及阳离子-π相互作用。突变结合口袋的相关氨基酸残基严重影响BetT对胆碱的转运。此外,关于胆碱的质子协同转运机制,作者通过功能实验找到了潜在的质子化位点D210,这一结果与结构上的发现相吻合 (图1)。
对于PsBetT的活性调控机制,作者根据结构分析提出了自抑制模型,并推测自抑制的活性调控机制涉及到跨膜结构域和C端结构域之间的相互作用。功能实验结果表明,野生型BetT仅在高盐条件下表现出较高的胆碱转运活性,而C端结构域截短体BetTΔCTD在低盐条件下就表现出较高的胆碱转运活性。C端结构域和跨膜结构域相互作用界面处的关键氨基酸残基的突变体同样在低盐条件下就表现出较高的胆碱转运活性。此外,自抑制机制依赖于BetT三聚体的形成,如果通过突变使其解聚成单体,则其单体在低盐条件下即可表现出较高的活性(图2)。
上述工作揭示了BetT的胆碱识别机制以及渗透压对其活性的调控机理,为理解同家族蛋白的活性调控机制提供了模板。此外,由于下游产物甜菜碱对植物抵御光抑制损伤具有重要的保护作用,该研究也为改造植物提高光合作用效率提供了重要的分子策略。
中国科学院分子植物科学卓越创新中心研究生杨天骄、博士后年宇薇、助理研究员林华建和武汉大学博士生李静为该论文的共同第一作者,中国科学院分子植物科学卓越创新中心范敏锐研究员,复旦大学张金儒研究员及爱荷华州立大学Gwyn A. Beattie教授为共同通讯作者。武汉大学王隆飞教授,复旦大学硕士生林湘及中国科学院分子植物科学卓越创新中心科研助理李天明、王睿瑛也参与了该研究。感谢中山大学刘建忠教授和华南理工大学罗立新教授提供大肠杆菌MKH13菌株。该研究得到中国科学院分子植物科学卓越创新中心、中国科学院先导科技专项和中国科学院上海分院的大力支持和资助。
论文链接:https://www.science.org/doi/10.1126/sciadv.ado6229
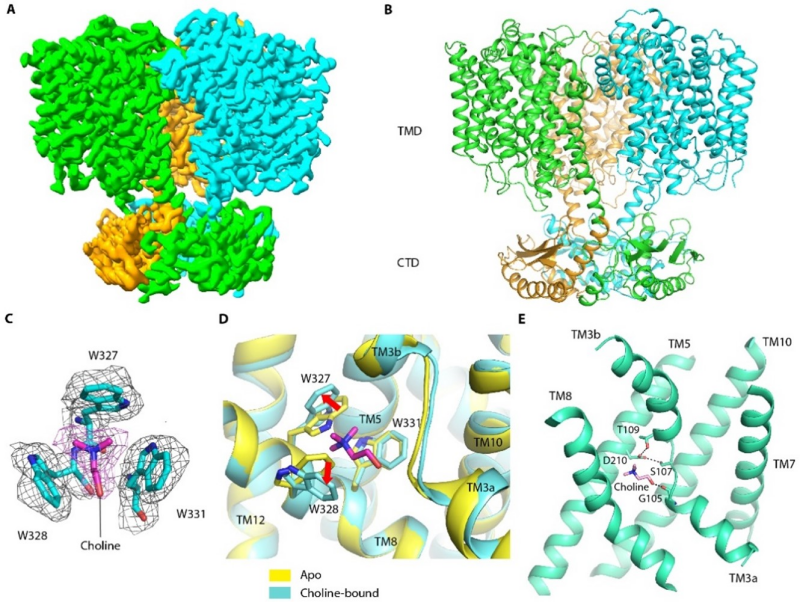
图1 PsBetT的整体结构以及底物识别与转运机制
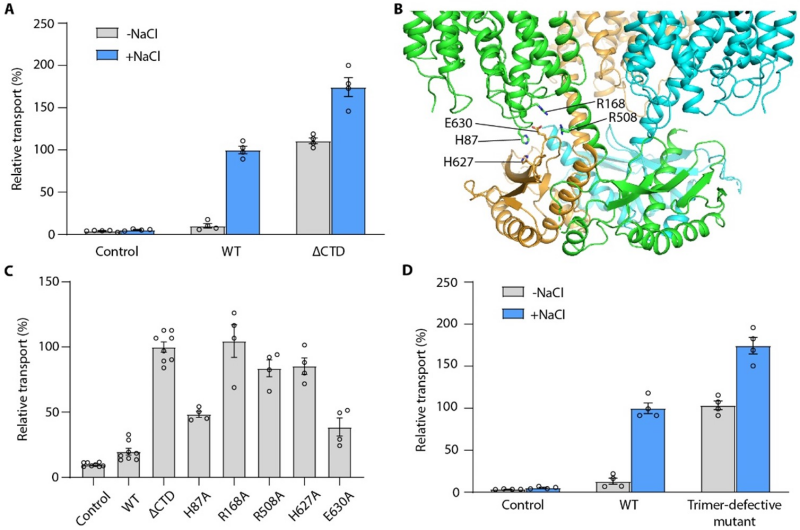
图2 PsBetT活性调控的自抑制机制
